
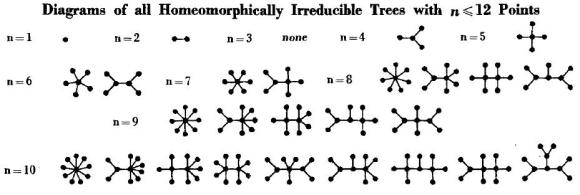
Esta escena de la película «El indomable Will Hunting» (título original «Good Will Hunting») de 1997, dirigida por Gus Van Sant y protagonizada por Matt Damon y Ben Affleck (en la que también aparece Robin Williams) describe un problema matemático de la teoría de grafos: dibujar un sistema de representantes de las 10 clases de árboles con 10 vértices homeomórficamente irreducibles. La solución de este problema fue obtenida por Frank Harary, Geert Prins, «The number of homeomorphically irreducible trees, and other species,» Acta Mathematica 101: 141-162, 1959 [copia gratis]. La solución a este problema para árboles con hasta 10 vértices aparece en la siguiente figura, extraída de dicho artículo (en formato un poco más compacto).
En teoría de grafos, un árbol es un grafo en el que cada par de vértices está conectado sólo por un único camino. Una definición más técnica nos dice que un grafo simple no dirigido G con un número finito n de vértices es un árbol si cumple cualquiera de las siguientes condiciones (todas equivalentes entre sí): (1) es conexo y tiene n-1 aristas, o (2) es conexo y no tiene ciclos, o (3) tiene n-1 aristas y no tiene ciclos, o (4) existe una única trayectoria entre cada par de vértices [gracias Alberto por el comentario de más abajo]. El grado de un vértice es el número de aristas a las que está conectado. Una «hoja» es un vértice de grado 1. Un vértice interno es un vértice de grado mayor que 1. Un árbol es irreducible si no tiene vértices de grado 2.
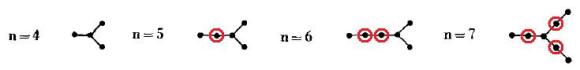
En esta figura, el único árbol irreducible es el de n=4, ya que los de n=5, 6 y 7 vértices todos se pueden reducir a dicho árbol eliminando los vértices de grado 2 que están marcados con una circunferencia en color rojo. Obtener todos los grafos con 10 vértices que son irreducibles no es difícil y cualquiera tanteando un poco puede hacerlo. Obviamente, lo más difícil es demostrar que no existe ningún otro.
¿Te atreves a dibujar todos los árboles irreducibles con n=11? No te pide que demuestres que tu lista es completa, sólo que lo intentes. No es un problema difícil (no creo que te requiera más de una hora de trabajo). Si lo haces te sentirás como Matt Damon (o como Will Hunting) Por supuesto, puedes chequear tu respuesta con el artículo original (que presenta los grafos irreducibles hasta n=12); también, hay programas de ordenador en la web que te permiten dibujarlos (si te gusta programar en Mathematica este problema es bonito para resolver por uno mismo sin buscar el notebook en la web). ¿Eres profesor de matemáticas? Por qué no le propones este problema a tus alumnos (resuelves el caso hasta n=6 y les pides a los alumnos que tanteen los casos n=7 u n=8).
Esta entrada participa en la Edición 4.1 del Carnaval de Matemáticas cuyo anfitrión es Tito Eliatron Dixit.


